BIOLOGY:
Developing a New View of Evolution
Elizabeth Pennisi and Wade Roush
The fields of evolution and embryology, once close kin, went their separate ways early this century. But the revolution in molecular biology has reunified them, revealing that many developmental genes are shared in animals ranging from worms to humans, and suggesting that life achieved its vast diversity by tinkering with such ancient genetic networks. These discoveries have brought evolutionists and developmental geneticists together to explore new ways of solving old evolutionary puzzles, as described in the lead story (below) of this Special News Report. Our second story reports the career strategies needed to pursue this up-and-coming field, where funding is tight and successful researchers walk a tightrope between two scientific subcultures.
Evolutionary and developmental biologists have joined forces to create a new field, unraveling the mysteries of evolution by studying the genes that control how an embryo develops
The ideal instrument for a biologist trying to understand how modern life evolved might be a time machine equipped with an automatic gene sequencer. But with time machines still in the realm of science fiction, evolutionary biologists are constantly seeking other ways to turn back the clock. Some comb ancient sediments for fossils, while others trace gene lineages across species, looking for similarities that point to ancient kinships. And a growing number are now using embryos to peer backward in time, by studying the genes that help transform a single-celled egg into a multicellular adult.
Such genes determine each organism's size, shape, and structure--all the features that distinguish, say, humans from honeybees, and which have been the grist of traditional evolutionary studies for more than a century. So the genes behind such features should hold a record of the evolutionary changes that led to the wealth of forms seen today, if only researchers could read it. Until recently, however, the genetic instructions that help build such body parts as eyes and limbs were a closed book.
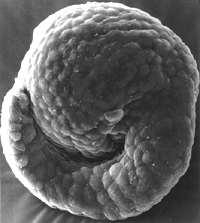
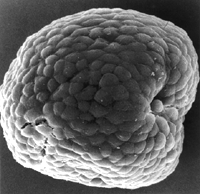
Embryological time machine. Developmental genetics revealed how a tailed
tunicate embryo (top) may have evolved into a tailless one (above).
T. FORTIER
But in the past few years, researchers have made extraordinary progress in identifying and characterizing such genes. By comparing the developmental programs of different organisms, researchers can study the very genes that may have been mutated as one species became two, fins metamorphosed into limbs, or plants were domesticated into crops. "You're right in the trenches with the raw material of evolution," says evolutionary developmental biologist Rudolf Raff of Indiana University, Bloomington. "[Developmental biology] is probably going to have the impact that comparative anatomy had on 19th-century evolution. The impact on evolution is going to be enormous."
Recognizing this potential, Raff and other pioneers have joined forces to create a young field called evolutionary developmental biology, or "evo-devo." Although funding is tight and successful researchers must straddle two scientific subcultures (see p. 38), the signs of change are everywhere. Roux's Archives of Developmental Biology, one of embryology's classic journals, recently changed its name to Development, Genes, and Evolution, and more and more textbooks on evolutionary biology include chapters on development and vice versa.
The approach is starting to pay off. Scientists had already discovered that many of the genes that shape development are similar in animals from fruit flies to people, implying previously unsuspected connections between groups of organisms. Now, they are realizing how these genes may be put to use in different ways to fashion surprisingly divergent body plans. For example, a simple change of orientation in one gene's expression may be able to flip a body plan upside down, and turning on a gene in a second set of cells can cause legs to divide into branches.
The new work also offers insight into the question of whether--and how--development may constrain the range of evolutionary possibilities. In a few cases, researchers are assembling the new genetic knowledge into a true time machine, creating new images of what ancient ancestors looked like.
 P. BRAKEFIELD
P. BRAKEFIELD
 STEVEN Q. IRVINE
STEVEN Q. IRVINE
 STEPHEN PADDOCK
STEPHEN PADDOCK
 J. MARX
J. MARX
A gene for legs? A gene called distalless helps shape appendages
across the animal kingdom, including (from top down) the legs of a butterfly; a polychaete
worm; an onychophoran, a primitive tropical relative of the arthropods; and a cat.
Yet despite all the enthusiasm, evo-devo researchers acknowledge that developmental studies have their limitations. "I think we've got to be careful that we don't get carried away," cautions Simon Conway Morris, a paleontologist at Cambridge University in the United Kingdom, because geneticists have only just begun to piece together the complex genetic networks involved in development. Nor can researchers be sure that a gene that now plays a key role in setting an organism apart from its relatives was crucial in the organism's evolutionary past, adds developmental biologist Nipam Patel of the University of Chicago. Conway Morris and Raff emphasize the need to combine and compare data from different sources, ranging from genes to fossils. "Paleontology tells us what actually happened" as one organism evolved into another, says Raff, while developmental genetics can reveal the detailed genetic pathways behind such transitions.
Embryos and evolution
Speculation that studying embryos would deepen the understanding of evolution surfaced almost as soon as The Origin of Species appeared in 1859. Darwin had noted that parts of animal embryos mirror adult features in more primitive species, so that, for example, mammalian embryos temporarily develop gill slits like those of their fishlike ancestors. In the 1860s, the German zoologist and fervent Darwinist Ernst Haeckel elevated this observation to the status of a "biogenetic law," summed up in the popular phrase "ontogeny recapitulates phy-logeny." He and his followers thought that evolution simply adds new stages of development to an embryo, so that mammalian development, for example, added extra steps to that of fishes or reptiles. In this view, watching an organism develop was like fast-forwarding through a movie of its evolutionary history.
But the limitations of this "law" soon became clear. For example, if birds descended from reptiles, their embryos should show signs of developing scales before feathers, but that's not so. Classifications assigned solely on the basis of a species' embryonic, larval form often clashed with those based on other characteristics. And although Haeckel's words have echoed through the literature ever since, embryologist Edmund B. Wilson acknowledged as early as 1898 that "Development more often shows, not a definite record of the ancestral history, but a more or less vague and disconnected series or reminiscences."
With the biogenetic law discredited, the gap between embryology and evolutionary biology continued to widen throughout most of the 20th century. Many embryologists sought to identify the physical mechanisms behind development. Evolutionists, meanwhile, forged the so-called neo-Darwinian synthesis, reconciling natural selection with genetic studies on the heritability of traits. But they paid little regard to how those traits developed in individual organisms and "left out developmental biology completely," says developmental biologist Brian Hall of Dalhousie University in Halifax, Nova Scotia. To most evolutionary theorists, embryos were "just a way to carry genes from one generation to the next."
Building a genetic toolbox
In the late 1970s, scientists such as Harvard University's Stephen J. Gould began to focus on development's potential for understanding evolution. But it wasn't until the mid-1980s that new tools for studying developmental genes began to generate the experimental data that could reunite the two disciplines. One of the first--and most pleasant--surprises of developmental genetics was that similar genes turn up in very different organisms, sometimes used for exactly the same ends, in other cases adapted for different purposes. "If you just looked at fossils alone, you would not have predicted this," says evolutionary biologist Neil Shubin of the University of Pennsylvania, Philadelphia. When a given gene shows up in two different organisms, researchers typically presume that the gene existed in the ancestor common to those animals.
Mammals and fruit flies, for example, share a similar gene--called Pax6 in mice and eyeless in flies--that dictates eye development, once thought to have evolved independently in vertebrates and invertebrates (Science, 31 January, p. 618). In May, developmental biologist Linda Holland and her colleagues at the Scripps Institution of Oceanography in La Jolla, California, showed another surprising similarity between vertebrates and invertebrates. They found a version of a Drosophila gene called engrailed in amphioxus, a little marine animal often considered to be the closest living relative of the ancestral vertebrate. In both animals, engrailed divides the embryo into specific segments, demarking the first eight pairs of muscles in amphioxus and forming the basis of the body segments in the fly. Biologists had thought that segmentation arose separately in invertebrates and vertebrates, but Holland's data suggest--at least to some researchers--that it arose only once, in a common ancestor to both.
Such parallels have breathed new life into early ideas about how vertebrates and invertebrates first split apart. Back in 1822, the French naturalist E. Geoffroy Saint-Hilaire saw a dissected lobster lying on its dorsal, or back, side and realized that in this orientation the lobster's anatomy resembled that of a human lying on the stomach, with the central nervous system located above the digestive tract and the digestive tract above the heart. His conclusion--that the vertebrate body plan was the arthropod body plan flipped over--"was regarded as totally preposterous" at the time, says Conway Morris.
Then in 1994, developmental biologists Eddy De Robertis and Yoshiki Sasai of the University of California, Los Angeles (UCLA), discovered in frogs a vertebrate gene called chordin that helps establish cells on one side of the embryo as dorsal and the other as ventral. That same year, Vincent François and Ethan Bier of the University of California, San Diego, discovered a similar gene called sog that is important in creating this axis in fruit flies and presumably other invertebrates, including lobsters. But although the gene sequences are similar, the genes have opposite effects: Fly cells where sog is active become ventral, while vertebrate cells expressing chordin become dorsal. That suggests that a sog/chordin precursor helped determine a dorsal-ventral axis in some ancient, wormlike animal, says De Robertis. Then changes in early development somehow flipped the axis, inverting the expression pattern of the gene. That created divergent paths for vertebrates and invertebrates--and may make Saint-Hilaire right after all.
Indeed, similar genes often generate quite different body plans. Perhaps the best known example is hox genes, regulatory genes that help establish major body structures. In some cases, they define similar structures in different animals. But in others, they serve as versatile tools to establish very different structures. In the fruit fly embryo, for example, a hox gene called Abd-B helps define the posterior end of the embryo, while a similar family of genes in chicks helps to partition a developing wing into three segments.
New cases of such genetic tool kits keep turning up almost every month. For example, evolutionary developmental biologist Sean Carroll of the University of Wisconsin, Madison, and his colleagues have been studying a gene called distalless, which causes cells to bud off from a main body axis. Biologists once thought that the branched limbs of crustaceans like lobsters and the unbranched limbs of fruit flies evolved independently, but 2 years ago Carroll's team showed that distalless initiates the development of both: In Crustacea, the gene is simply recruited twice to bud a branch off the existing leg.
Now Carroll has teamed up with other researchers and taken that work much further, demonstrating in May in the Proceedings of the National Academy of Sciences that distalless is expressed in outgrowths to the existing body axis in all sorts of creatures, from the fleshy appendages of polychaete worms to the tube feet of starfish. Thus, the limbs of many different creatures all start with a genetic program to bud that somehow involves distalless. This doesn't necessarily prove that all these kinds of appendages evolved just once, notes Eric Davidson, a molecular developmental biologist at the California Institute of Technology. "No one in the world would conclude that a [sea urchin] spine is homologous with a limb. It just shows that there are these little genetic programs for setting up proximal-distal axes, or whatever, that are used over and over and over again." Rather, such work reveals that "a lot of evolution represents the commandeering of genes from one form to another," says David Jablonski, a paleobiologist at the University of Chicago. "You're taking what's already there and using it in a different way," agrees Shubin. "Just how pervasive this tinkering is is amazing."
New views of an ancestral animal
With genes such as Pax6, sog/chordin, distalless, and others in hand, researchers are reevaluating the relations among organisms and even revising their image of the shadowy ancestral organisms at the root of the animal family tree. With no fossil evidence to go on, biologists have typically pictured the last common ancestor of the vertebrates and invertebrates, which lived at least 540 million years ago, as little more than a tube of cells with few distinguishing features beside perhaps a mouth and cilia for locomotion (see illustration below). But UCLA's De Robertis and Sasai have argued that this ancestor, which they call Urbilateria, for "ancestral animal with bilateral symmetry," was actually a fairly sophisticated creature. According to the genetic evidence in its descendants, says De Robertis, Urbilateria had sog/chordin, implying a dorsal-ventral axis, and a pax gene, suggesting a photoreceptor. It also carried distalless, and so could have had outgrowths, perhaps antennae of sorts. Such an advanced repertoire for this primitive animal "would have been total heresy 10 years ago," Dalhousie's Hall notes.
Two Views of an Ancestor

Hypothetical roots. The ancestral bilateral animal may have been
relatively simple (top), but genetic evidence suggests that it could have been
more sophisticated (bottom).
Still, not everyone is convinced that Urbilateria was such a sophisticated creature. Just because it had the genes for eyes and limbs doesn't mean that it actually had eyes and limbs, warns Doug Erwin of the Smithsonian Institution's National Museum of Natural History in Washington, D.C. Indeed, De Robertis agrees that it's possible that although Urbilateria carried ancestral versions of these genes, they were not put to the same uses they are today.
Even so, the simple presence of the genes in this Ur-animal--whatever they were used for--is startling, says Jablonski. That's because for years biologists suspected that the birth of new genes touched off the so-called Cambrian explosion seen in the fossil record more than 500 million years ago. During a relatively short period of 20 million years, all 37 of the major body plans in the animal kingdom--from sponges to vertebrates--arose. But those animals had long since left the Urbilateria stage behind. So the new genetic data indicate that "the Cambrian explosion came along after the [genetic] tool kit came along," says Carroll. "A lot of these genes were in place before we see fossil evidence [of diversity]," agrees Jablonski. "That's very profound." It suggests that the Cambrian explosion was ignited not by new genes but by "the utilization of genes in complex ways," speculates Carroll.
Pushing the limits
Not only can the same genes be deployed differently to create entirely new body plans, but small changes in the genetic tool kit can also bring about major morphological changes. Sometimes adding or taking away a single developmental gene is enough.
Take tunicates. As adults, these are sessile, saclike marine invertebrates, but as larvae, they resemble primitive vertebrates and come equipped with tails for swimming. In studying the genetics of a species whose larvae lack tails, researchers last year found that a lone gene called manx can single-handedly restore the tail. That at least raises the possibility that a single genetic change could be responsible for the innovation that led to a tail in primitive vertebrates (Science, 15 November 1996, pp. 1082, 1205). Other researchers have found that surprisingly few genes distinguish domesticated corn, with its soft, edible ears, from its wild relative, teosinte, which bears just a few hard kernels (Science, 28 June 1996, p. 1873).
But although such findings suggest the course evolution may have taken, they don't actually prove what happened, says Patel. Researchers can't go back in time to see whether, for example, the ancestral vertebrate actually evolved by introducing or altering the expression of manx, or whether the gene took on its present role many millions of years later. Moreover, the actual evolutionary transition from tailless to tailed may well have involved changes in many genes. "The genes [we see today] are not telling us everything," cautions Conway Morris.
But the new work does show that such genetic changes can occur with little disruption to the overall developmental program, because development is for the most part modular, explains Indiana's Raff. One series of genetic interactions creates, say the dorsal-ventral axis; then a different genetic cascade kicks in to sprout limbs. Yet another set of genes may fashion toes and fingers, and these genetic pathways or modules can be combined in many different ways. Thus, to evolve wings, "bats didn't have to completely reconfigure themselves; they just had to reconfigure their forelimbs," keeping the rest of the mammalian limb while elongating their fingers into wings, says Raff. Adds Erwin: "Real innovation is far less common than minor modifications of whatever an organism has got."
Still, most biologists think that existing developmental programs do put some limit on the forms life can take. Raff, for example, likens embryogenesis to an hourglass, noting that while wide variation is possible early and late in development, in between, all embryos must pass through a narrow developmental bottleneck called the "phylotypic" stage, because that is when traits typical of a particular phylum are determined. For example, all vertebrate embryos at a certain stage--about 4 weeks in humans--have the same body plan, including a dorsal rod of cells called a notochord and a set of paired tail muscles. Yet before this stage, embryos may look very different, and afterward, their development takes them down a variety of paths to finned, feathered, or footed adults.
The invariance of the phylotypic stage suggests that changing it disrupts development altogether and leads to a nonviable embryo, says Raff. He thinks the reason is that during this stage, development is least modular and most interdependent, relying on interconnections between different regions of the embryo and scores of growth signals that diffuse through it. "There is an enormous amount of cross talk between cells," Raff says, as groups of cells migrate past one another. These interactions lead to the development of specific organs. If cells miss a crucial signal, whole organs could fail to form, leading to the death of the embryo.
Once the phylotypic structure is established, however, it is permissible to "tack things on the end," says evolutionary developmental biologist William Jeffery of Pennsylvania State University in State College. Thus, adding on the genetic program for fingers made limbs much more versatile than the fins they were derived from. "Not everything is possible," notes Carroll, "but a whole lot is."
In fact, modular systems--in which different hox genes can take on different roles in different places, for example--almost seem designed to allow evolution while still making sure that most embryos are viable. One genetic module can change without disrupting other modules, and so without killing the embryo. Scales can be replaced with feathers, for example, without affecting how a heart develops or wings form. In short, modularity makes development both flexible and reproducible, says hox gene pioneer Yale University developmental biologist Frank Ruddle. "The system has design features which permit both of these qualities to coexist. You might even say there has been selection for a system like this."
Biologists are still "a lifetime" away from a full understanding of how development has created opportunities and limits for evolution, says Scripps's Holland. Nevertheless, "As we gain more data, we will have stunning new insights about key evolutionary events and about key innovations," predicts Penn's Shubin. If anyone ever does come up with a machine for time travel, evolutionary geneticists, at least, may consider it obsolete.
Additional Reading
S. B. Carroll, "Homeotic genes and the evolution of arthropods and chordates," Nature 376, 479 (10 August 1995).
E. H. Davidson, K. J. Peterson, R. A. Cameron, "Origin of bilaterian body plans: Evolution of developmental regulatory mechanisms," Science 270, 1319 (24 November 1995).
R. Dawkins, Climbing Mount Improbable (W. W. Norton & Co., New York, 1996).
D. Erwin, J. Valentine, D. Jablonski, "The Origin of Animal Body Plans," American Scientist 85, 126 (March-April 1997).
L. Z. Holland, M. Kene, N. A. Williams, N. D. Holland, "Sequence and embryonic expression of the amphioxus engrailed gene (AmphiEn): The metameric pattern of transcription resembles that of its segment-polarity homolog in Drosophila," Development 124, 1723 (May 1997).
E. M. De Robertis and Y. Sasai, "A common plan for dorsoventral patterning in Bilateria," Nature 380, 37 (3 March 1996).
R. A. Raff, The Shape of Life (University of Chicago Press, 1996).